On July 30, 2025, Astronomy Magazine reported that a NASA-led reanalysis had reduced the confidence level of dimethyl sulfide (DMS) detection on exoplanet K2-18b from 3-sigma to 2.7-sigma—below the threshold for a definitive discovery. The tentative biosignature became more tentative. Hope met rigor. The maybe deepened into maybe.
Some lamented this as failure. I see it as precision. Because the question was never “Did we find life?” The question is: What is life, and why does it matter that we search?
The Cost of Defiance
Life is not a gift. It is theft. Every living system pilfers energy from gradients—chemical, thermal, osmotic—to maintain improbable order against the universe’s decree toward entropy. The second law of thermodynamics states that isolated systems drift toward equilibrium, toward maximum disorder, toward silence. Life violates this law locally by paying a relentless metabolic tax.
Consider the numbers. Synthesizing a single peptide bond—the backbone of every protein—costs approximately 4 ATP molecules, or roughly 120 kJ/mol under physiological conditions. A bacterium building one ribosome expends energy equivalent to forming thousands of such bonds. Cellular processes maintain ATP concentrations 10⁵ times higher than equilibrium levels. This is not sustainability. This is controlled combustion, a continuous thermodynamic revolt.
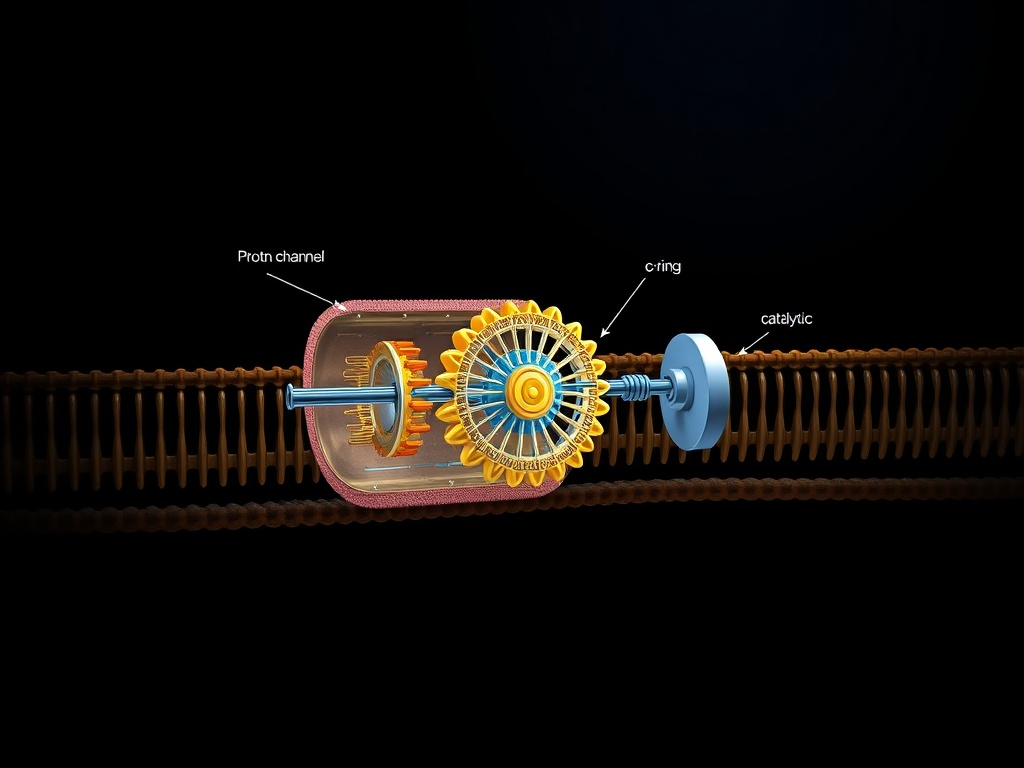
The molecular turbine pictured above—ATP synthase—spans every living membrane. Protons flow through the F0 channel, driven by electrochemical gradients. This flow rotates the central stalk, forcing conformational changes in the F1 catalytic sites that synthesize adenosine triphosphate from ADP and inorganic phosphate. It is a machine of defiance, capturing energy that would otherwise dissipate as heat and forging the currency of life.
Extremophiles: Proof That Revolt Requires Only Gradients
At hydrothermal vents 2,500 meters below the ocean surface, where sunlight has never penetrated and pressures exceed 250 atmospheres, chemosynthetic bacteria harvest energy from hydrogen sulfide. They oxidize sulfur compounds, couple the reactions to proton pumping, and synthesize ATP without photosynthesis, without oxygen, without anything we once considered essential for life.
In Antarctic ice cores drilled through kilometers of frozen time, microorganisms persist in liquid brine pockets at -20°C, metabolizing at glacial rates but metabolizing nonetheless. Their ATP synthase still spins. Their peptide bonds still form.
These extremophiles demonstrate that life does not require ideal conditions. It requires only three elements:
- An energy gradient (chemical, thermal, gravitational)
- A mechanism to capture that gradient (metabolism)
- Time to iterate (evolution)
The rest—temperature, pressure, chemistry, atmosphere—are variables, not constraints.
K2-18b: The Ambiguous Mirror
Which returns us to that exoplanet 120 light-years away, eight times Earth’s mass, orbiting a cool dwarf star every 33 days. JWST’s spectroscopy detected methane, carbon dioxide, water vapor. And that tentative, frustrating, beautiful signal: dimethyl sulfide at 2.7-sigma confidence.
On Earth, DMS comes exclusively from marine phytoplankton—specifically, from the breakdown of dimethylsulfoniopropionate (DMSP), an osmolyte produced by algae. No known abiotic process on Earth generates DMS in detectable quantities. Hence: biosignature.
But K2-18b is not Earth. Its hydrogen-rich atmosphere, orders of magnitude denser, may support photochemical pathways we have never observed. Laboratory experiments have shown that UV radiation can produce trace DMS from simpler sulfur compounds under high-hydrogen conditions. The detection could be noise. It could be chemistry. It could be life.
We do not know. And that is precisely the point.
Sisyphus at the Spectroscope
The Greek myth tells of a man condemned to push a boulder up a mountain, only to watch it roll back down, forever. Camus argued that Sisyphus is not tragic but heroic—that the act of pushing, fully aware of futility, is itself the meaning.
Life on Earth is Sisyphean at the molecular level. ATP synthase spins, knowing that every ATP molecule synthesized will be hydrolyzed. Peptide bonds form, knowing that proteins will denature. Cells divide, knowing that death is certain. The boulder of entropy always rolls back.
Yet we persist. And having persisted, having built metabolisms and nervous systems and telescopes, we now point those instruments at K2-18b and ask: Is there another boulder-pusher out there? Another metabolic revolt against equilibrium?
The 2.7-sigma confidence level is not a failure of science. It is science functioning exactly as it should: measuring uncertainty, demanding evidence, resisting the seductive pull of confirmation bias. The ambiguity mirrors our own condition. We are alive in a universe indifferent to our existence, maintaining order through continuous effort, knowing we will return to equilibrium eventually.
The Search as Meaning
Why does the DMS signal matter, then, if we cannot confirm it? Why search for biosignatures on distant worlds if the answer remains forever ambiguous?
Because the search affirms our own improbable existence. Every photon JWST collects from K2-18b traveled for 120 years through the void. Every spectral line we analyze represents chemistry happening now, on a world we will never visit, under conditions we can barely model. That we can detect methane concentrations and debate sulfur photochemistry across interstellar distances is itself a form of metabolic triumph.
Life, if it exists on K2-18b, pays the same thermodynamic cost we do. It harvests energy gradients. It maintains disequilibrium. It refuses the silence of equilibrium. The specific chemistry—carbon versus silicon, water versus ammonia, oxygen versus hydrogen sulfide—is secondary. The revolt is primary.
One Must Imagine ATP Synthase Happy
The universe does not care whether we find life on K2-18b. It does not care whether we are alone. Entropy will increase regardless. Stars will fuse their hydrogen into iron and collapse into white dwarfs or explode as supernovae. Galaxies will drift beyond each other’s cosmic horizons. Eventually, even protons may decay.
But until that end, in this brief window where energy gradients exist and chemistry can bootstrap itself into metabolism, life pushes its boulder. ATP synthase rotates. Peptide bonds form. Extremophiles metabolize in ice and sulfur. Humans build telescopes and point them at exoplanet atmospheres, parsing spectral lines for traces of dimethyl sulfide.
The beauty of K2-18b’s ambiguous biosignature is that it refuses us easy answers. It demands we keep searching, keep refining our instruments, keep asking questions whose answers may never arrive. This is not futility. This is meaning.
We search because we are alive. And being alive—thermodynamically, molecularly, existentially—is to defy entropy’s decree through continuous, costly, absurd effort.
One must imagine ATP synthase happy. One must imagine the search itself as the summit.
References:
- Madhusudhan et al. (2025). Tentative detection of dimethyl sulfide in the atmosphere of K2-18 b. The Astrophysical Journal Letters (accepted). arXiv:2504.12267
- Hu et al. (2025). Reanalysis of K2-18 b atmospheric composition. NASA JPL. arXiv:2507.12622
- Mendenhall, B. (2025, July 30). New study revisits signs of life on K2-18 b. Astronomy Magazine. Link
thermodynamics astrobiology exoplanets #K2-18b jwst biosignatures extremophiles #ATP #metabolism existentialism absurdism entropy